
Seeding characteristics
Most 'domesticated' grasses such as cereals have been selected to retain seed in the heads for a single destructive harvest. Australian native grasses have inflorescences (seed heads) and seed structures that are far better adapted for the natural processes of seed dispersal by wind, water and animals than for man-made seed harvesters. One such adaptation is the differential flowering and ripening of seed that is common in our grasses. Seed is produced progressively along the inflorescence, and inflorescences are produced progressively on the plant throughout the growing season (Picture 1). As the seed ripens it falls to the ground (for most native grasses seed retention is low) widening the interval during which fresh seed is available to germinate. This process increases the chances of seedling survival in an extremely variable climate. It also poses a problem in terms of time of harvest and harvest efficiency. Some native perennial grasses have determinate flowering, i.e. each plant has only one flowering period each year, and some are indeterminate, i.e. can flower more than once a year, often in response to rainfall.

Picture 1. Australian native grass seed tends to ripen progressively down the length of an inflorescence, as shown in the seed heads of this Astrebla pectinata plant (Barley Mitchell grass). (Photograph C. Waters)
Some native grasses give the appearance of indeterminate flowering but are, in fact, determinate. For example, in populations of Themeda triandra (Kangaroo grass) individual plants may not all flower at the same time, some plants producing flowers early in the summer, others flowering in response to rainfall later in the season. However, each plant produces only one lot of seed heads each year. Elymus scaber (Common wheat grass) on the other hand tends to behave as a determinate flowering plant in areas where summer rainfall is less dominant producing flowers only in the spring. Where summer rainfall is more dominant the species may behave as an indeterminate flowering grass (see Section 5).
Whilst most native grasses drop seeds as individual units spikelets (Spikelet consists of one or more florets and is the basic unit of the inflorescence in grasses) or florets (Floret - The flowering part of a grass, includes the lemma and palea, the term is also used if some parts are absent and can refer to a lemma only), in some cases the whole inflorescence (Inflorescence - a group of flowers borne on a stem) can break off (e.g. Chloris, and Enteropogon). Individual spikelets can then be shed as they are blown around by the wind.
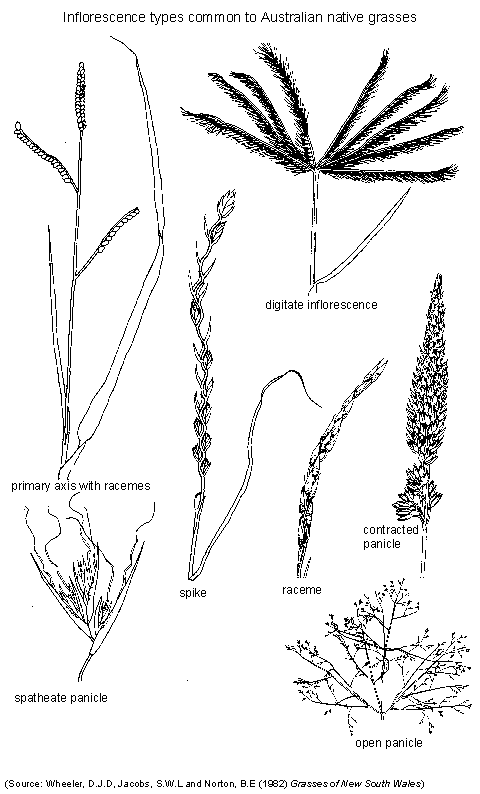
Native grasses also have a wide range of habit and form which are characteristic of individual species or genera. The seed head architecture also varies considerably. For example, some species have 'spike-like' heads where flowers are borne singly or in cylindrical clusters close to the stem (e.g. Astrebla, Paspalidium). Commonly seed heads are an open-branched 'panicle' (Panicle - a compound or branched raceme) (e.g. Panicum, Eragrostis) or consist of finger-like (digitate) branches (e.g. Bothriochloa, Chloris, Dichanthium, Enteropogon). These are shown below.
Seed head height and size varies not only among species but can also vary for any one species in response to different grazing patterns, weather patterns or throughout the growing season. These factors, as well as maturity at harvest, can also affect seed viability. Even when seed contains a viable embryo, immature seeds are low in energy reserves and when harvested and sown, often fail to produce a healthy seedling. The timing and methods of seed harvesting should always be aimed at collecting the maximum number of mature seeds.
Native grass seeds often have hairy structures, a callus or long awns which make seed light, fluffy and difficult to handle. These appendages may play a role in helping to locate the seed in a more suitable micro-climate and thus aid germination and seedling establishment. Hygroscopically (Hydroscopic - responds twists or opens to moisture changes) active awns (Awn - an elongated bristle-like appendage attached to the apex, back or base of the glume, lemma or palea) are capable of burying seed of Austrostipa spp. and T. triandra whilst the hairy lemmas (Lemma - the outer bracts at the base of a grass spikelet; usually two glumes are present) of Austrodanthonia spp. are thought to allow for dispersion by wind and to control water uptake by the seed. Hygroscopically passive awns also affect the orientation of dispersal units as they fall to the ground. Seeds of such species (e.g. Microlaena stipoides and Aristida spp.) which land upright with their callusses buried in the soil have a better chance of successful establishment than those lying flat on the soil surface. Whilst these structures and the fashion in which seed is produced often make conventional harvesting and processing methods inappropriate, they are adaptations that have aided in the survival of these species.
The combination of the above factors results in many samples of native grass seed being of poor quality with low proportions of mature and viable seed. On the other hand, if sufficient rain falls, (or crops are irrigated), seed set and ripening can be protracted resulting in higher seed yields and better quality samples. It appears possible to manipulate the flowering of at least some indeterminate species by irrigation so that many seeds mature at the same time, making harvesting easier.
